REGULACIÓN DIFERENCIAL DEL EJE HIPOTÁLAMOHIPÓFISISTIROIDEO EN LA ANOREXIA Y
ley Orgánica 32021 de 24 de Marzo de RegulaciónMemoria de Buena Regulación Normativa y Económica Sobre el
INTRODUCCIÓN

Regulación diferencial del eje hipotálamo-hipófisis-tiroideo en la anorexia y la restricción alimentaria en la rata
a,bLorraine Jaimes, a,bPatricia de Gortari*, cPatricia Joseph-Bravo
aInstituto Nacional de Psiquiatria RFM, bEscuela de Dietética y Nutrición ISSSTE, [email protected]
cInstituto de Biotecnología, UNAM. México
RESÚMEN
El TRH, sintetizado en el núcleo paraventricular del hipotálamo (NPV), regula el eje tiroideo. Su metabolismo es regulado por el estado de nutrición. Para estudiar la participación del TRH en la conducta alimentaria se utilizó el modelo de anorexia por deshidratación (AN) y un modelo de restricción alimentaria (RA), induciendo una disminución de peso similar; y se utilizó el modelo de estrés por frío para determinar si los cambios en la AN son específicos al modelo, ya que las neuronas TRHérgicas del NPV se activan durante el frío. Se semicuantificó el RNAm de pro-TRH y TRH-R1 por RT-PCR del NPV. La AN y la RA disminuyeron su consumo de alimento y perdieron peso; la concentración sérica de leptina disminuyó y la de corticosterona aumentó. En el NPV de la RA, el RNAm de proTRH y de TRH-R1 disminuyeron; TSH y las hormonas tiroideas disminuyeron. El estímulo del frío activó las neuronas TRHérgicas en la RA, reflejado por un aumento del RNAm de proTRH. En cambio en la AN la expresión de proTRH aumentó en el NPV, al igual que la concentración sérica de TSH y T4. El estímulo del frío no provocó una mayor activación de las neuronas TRHérgicas.
INTRODUCCIÓN
La anorexia nervosa es un complejo trastorno alimenticio que se caracteriza por alteraciones de la conducta alimentaria, rechazo a mantener el peso corporal mínimo normal, miedo intenso a ganar peso y alteración en la percepción de la imagen corporal. La anorexia está asociada a una serie de alteraciones neuroendócrinas. El eje hipotálamo-hipófisis adrenal (HHA) está activado y el gonadal (HHG) se encuentra inhibido, en cuanto al eje tiroideo (HHT), existe controversia.
El eje HHT está regulado por la hormona liberadora de tirotropina (TRH) sintetizado en el Núcleo paraventricular del hipotálamo (NPV); es liberado de la eminencia media, y estimula la síntesis y liberación de tirotropina (TSH) en la adenohipófisis, ésta a su vez estimula la síntesis y liberación de las hormonas tiroideas (T3 y T4) en la glándula tiroides, las cuales por retroalimentación negativa inhiben la síntesis de TSH y de TRH en la adenohipófisis y en el NPV, respectivamente (Nillni y Sevarino, 1999; Yen, 2001). El TRH se libera en respuesta a estímulos ambientales como el frío, activándose el eje tiroideo. El eje HHT regula el metabolismo energético y la temperatura corporal (Griffihs, 1985; Leibel y Rosenbaum, 1995).
El TRH puede actuar como un neuropéptido anorexigénico (Nillni y Sevarino, 1999; Horita, 1998). Su metabolismo es regulado por el estado de nutrición; en el ayuno, el eje tiroideo se inhibe, y el contenido de TRH y los niveles de pro-TRH RNAm en el NPV disminuyen, mientras que en la eminencia media aumenta el contenido del péptido, reflejando una inhibición en su liberación que coincide con una disminución de la concentración sérica de TSH y de las hormonas tiroideas; paradójicamente la retroalimentación negativa está bloqueada (Blake et al., 1991; Van Haasteren et al., 1995; de Gortari et al., 2000).
La leptina es una hormona que juega un papel clave en la regulación del eje tiroideo en la restricción alimentaria. Esta hormona sintetizada en el tejido adiposo regula positivamente la expresión del gen de TRH en el NPV. La administración de leptina a roedores en ayuno, desinhibe el eje tiroideo, es decir, aumenta la concentración de las hormonas tiroideas y la expresión de TRH en el NPV (Legradi et al., 1997; Harris et al., 2001; Kim et al., 2000; Nillni et al., 2000; Guo et al., 2004).
El objetivo de este estudio es definir la participación del TRH hipotalámico en el control central de la conducta alimentaria y determinar si la activación del eje HHT que se observa en las ratas con anorexia es específica al modelo de anorexia inducida por deshidratación.
METODOLOGÍA
Se utilizó el modelo de anorexia inducida por deshidratación (restricción voluntaria del consumo de alimento) y un modelo de restricción alimentaria (restricción forzada, pareado al grupo de la anorexia), induciendo una disminución de peso similar. Además, para determinar si los cambios observados en el eje HHT en las ratas con anorexia son específicos al modelo, se expusieron a la mitad de las ratas de cada grupo al estímulo del frío (4 °C) durante una hora, estímulo que activa las neuronas TRHérgicas del NPV. Los animales fueron sacrificadas inmediatamente después de haber transcurrido la hora.
Se usaron ratas Wistar hembras adultas (230 g de peso), se mantuvieron en cajas individuales para registrar su consumo de alimento, agua y peso diariamente. El experimento tuvo una duración de siete días.
Los grupos de estudio fueron los siguientes: Control, n=10: con consumo de alimento y agua ad libitum; Anorexia por deshidratación (AN), n=11: con consumo de alimento ad libitum y una solución salina al 2.5% como bebida (Watts et al., 1999); Restricción Alimentaria (RA), n=10: con consumo de agua ad libitum y alimento en la misma cantidad que consumía el grupo de la anorexia.
Todos los animales se sacrificaron al octavo día de iniciado el experimento por decapitación. Se colectó la sangre de cada rata para obtener el suero. Se extrajeron los cerebros y se almacenaron a -70°C hasta su disección. La grasa subescapular y la epididimal fueron disecadas en fresco y se pesaron inmediatamente. El NPV se disecó manualmente utilizando un atlas de cerebro de rata (Palkovits y Brownstein, 1988). Se extrajo el RNA por el método de Tiocinato de guanidina (Chomczynski y Sacchi, 1987) y posteriormente se semicuantificó el RNAm de pro-TRH y TRH-R1 (receptor de TRH) por RT-PCR. En suero se midió la concentración de leptina por el método de ELISA; y corticosterona, hormonas tiroideas, TSH y estrógenos por radioinmunoensayo.
Se obtuvo para todos los datos la media y el error estándar de la media (ESM). Para realizar comparaciones entre los grupos se usó un análisis de varianza unidireccional (ANOVA) seguido por la prueba post-hoc de Fisher (software Stat View, SAS Institute). Se consideró una diferencia estadísticamente significativa cuando la p<0.05. Para analizar los datos del experimento del frío se utilizó un ANOVA de dos vías.
RESULTADOS
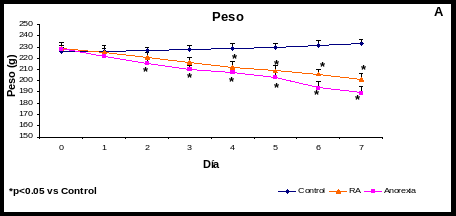
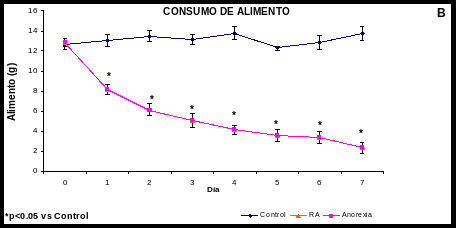
Cambios en el Peso, en el consumo de alimento y agua (Gráfica 1)
La RA perdió peso significativamente a partir del 4to día (205 4 g vs 223 6 g del control; p<0.05); el último día del experimento tuvo una pérdida de peso de un 17% vs el control (p<0.05) y disminuyó su consumo de agua en un 10% sin ser significativo.
En la AN las ratas disminuyeron su consumo de alimento a partir del primer día (10 0.6 g vs 17 1 g vs control; p<0.05), para el séptimo día disminuyeron su consumo de alimento en un 87% vs el control; p<0.05. Así mismo bajaron de peso significativamente desde el segundo día (204 4 g vs 222 6 g del control; p<0.05); para el séptimo día el grupo de la anorexia perdió un 25% de peso vs el control; p<0.05; y aumentaron significativamente su consumo de líquido en un 61% vs el control; p<0.05.
El peso de la grasa blanca (WAT) disminuyó tanto en la RA como en la AN con respecto al control (RA: 16 2%; AN: 17 1% vs 22 ± 2% del control; p<0.05). El peso de la grasa parda (BAT) mostró una tendencia a disminuir, sin embargo no fue significativa (Tabla 2).
Gráfica 1.
Cambios en leptina, corticosterona y estrógenos (Tabla 2)
La concentración de leptina disminuyó tanto en la RA como en la AN un 68% y un 64% respectivamente vs el control (100%; p<0.05). No se encontraron diferencias significativas entre la RA y la AN.
La concentración sérica de corticosterona aumentó en la RA y en la AN un 206% y un 168% respectivamente vs el control (100%; p<0.05). No se encontraron diferencias significativas entre la RA y la AN.
La concentración sérica de estrógenos disminuyó un 64% en la RA y un 70% en la AN vs el control (100%; p<0.05).
|
|
Control |
RA |
AN |
|
Leptina (pg/ml) |
422.05 36.67 |
136.25 14.74 a |
151.07 23.41 a |
|
Corticosterona (pg/ml) |
411.18 169.03 a |
282.13 22.30 a, b |
|
Estradiol (pg/ml) |
30 9 |
11.00 3 a |
9.00 3 a |
|
WAT (g/kg b.w.) |
21.94 1.71 |
16.34 1.67 a |
16.74 1.24 a |
|
BAT (g/kg b.w.) |
1.89 0.26 |
1.43 0.13 |
1.46 0.16 |
Table 1. a vs C, b vs FR
Cambios en el eje Hipotálamo-Hipófisis-Tiroideo
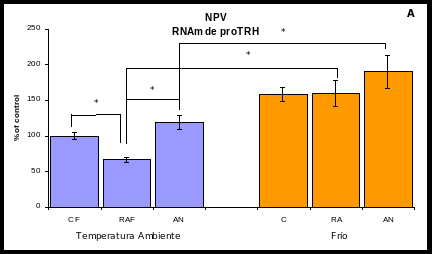
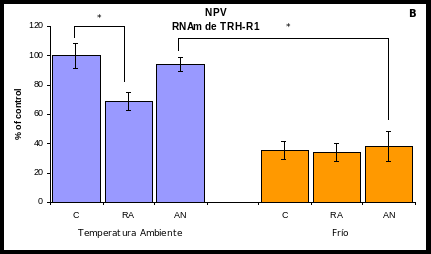
El eje HHT en la RA se encontró inhibido: la expresión de pro-TRH (Gráfica 2A) y la de TRH-R1 (Gráfica 2B) en el NPV disminuyeron un 38% (100 7 % del control; p<0.05)
La concentración sérica de TSH disminuyó un 40% con respecto al control (p<0.05). La concentración sérica de T3 y T4 disminuyeron un 30% y un 49% respectivamente vs el control (p<0.05) (Tabla 2).
Sin embargo, en la AN no se presentaron las mismas adaptaciones neuroendócrinas que en la RA a pesar de que la disminución de peso y el consumo de calorías y proteínas fue el mismo. En la AN la expresión de TRH en el NPV aumentó un 60% (100 7 % del control; p<0.05) (Gráfica 2A). La expresión de TRH-R1 no cambió significativamente (Gráfica 2B).
La concentración sérica de TSH aumentó un 44% con respecto al control y un 85% con respecto a la RA (p<0.05). Mientras que la concentración sérica de T3 disminuyó un 27% con respecto al control (p<0.05); en cambio T4 aumentó un 202% con respecto al control (p<0.05). Este aumento de T4 en la AN también fue significativo con respecto a la RA (p<0.05) (Tabla 2).
|
|
TSH (ng/ml) |
T3 (ng/dl) |
T4 (g/dl) |
Control |
2.11 0.22 |
73.12 4.7 |
1.94 0.27 |
|
RA |
1.07 0.09 a |
51.09 3.93 a |
0.99 0.11 |
|
AN |
3.05 0.39 a,b |
53.61 2.6 a |
5.87 0.59 a,b |
Tabla 2. a vs control, b vs RA
Gráfica 2.
Exposición al frío (Gráfica 2A y B): Como se mencionó anteriormente, el frío es una condición ambiental que estimula la activación de las neuronas TRHérgicas en el NPV. En el grupo control sometido al frío (CF) se encontró un aumento del 58% en la expresión de TRH en el NPV con respecto al control de temperatura ambiente (TA) (CF: 158 9% vs 100 7% del control; p<0.05).
El estímulo del frío provocó un aumento en la expresión de TRH en el grupo de la RA sometido 4 °C (RAF), a pesar de la restricción calórica, los niveles del RNAm de TRH aumentaron un 98% con respecto al grupo RA de TA (RAF: 160 18 % vs RA: 62 3 %; p<0.05).
En el grupo de la AN sometido al frío (ANF) no se encontraron cambios significativos con respecto al grupo de la AN de TA (ANF: 179 23% vs AN: 160 10%).
El estímulo del frío inhibió significativamente la expresión de TRH-R1 en el NPV: la expresión de TRH-R1 disminuyó en el grupo CF a un 35%; p<0.05, en la RAF disminuyó a un 34%; p<0.05 y en la ANF disminuyó a un 38%; p<0.05.
CONCLUSIÓN
A pesar de que se presentaron en la RA y la AN los mismos cambios metabólicos (disminución de la concentración de leptina y aumento de la concentración de corticosterona), y una disminución en el consumo de alimento, el eje HHT en la AN no presentó la misma adaptación que en la RA.
La falta de respuesta de las neuronas TRHérgicas del NPV en el grupo de la AN expuestas al frío sugieren que el cambio observado en la AN en el NPV es específico al modelo.
BIBLIOGRAFÍA
Blake NG, Eckland DJA, Foster OJF, Lightman SL. Inhibition of hypothalamic thyrotropin-releasing hormone messenger ribonucleic acid during food deprivation. Endocrinology 1991; 129:2714-2718.
Chomczynski P y Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 1987; 162:156-159.
De Gortari P, González-Alzati ME, Cisneros M, Joseph-Bravo P. Effect of fasting on the content of thyrotropin-releasing hormone and its mRNA in the central nervous system and pyroglutamyl peptidase II activity in the anterior pituitary of post-weaned and adult rats. Nutr Neurosci 2000; 3:255-265.
Griffiths EC. Thyrotropin releasing hormone: endocrine and central effects. Psychoneuroendocrinology 1985; 10:225-235.
Guo F, Bakal K, Minokoshi Y, Anthony N. Leptin signaling targets the Thyrotropin-releasing hormone gene promoter in vivo. Endocrinology 2004; 145(5):2221-2227.
Harris M, Aschkenasi C, Elias CF y cols. Transcriptional regulation of the thyrotropin-releasing hormone gene by leptin and melanocortin signaling. J. Clin. Invest. 2001; 107:111-120.
Horita A. An update on the CNS actions of TRH and its analogs. Life Sci 1998; 62:1443-1448.
Kim MS, Small CJ, Stanley SA y cols. The central melanocortin system affects the hypothalamo-pituitary thyroid axis and may mediate the effect of leptin. J. Clin. Invest. 2000; 105: 1005-1011.
Legradi G, Emerson CH, Ahima RS, Flier JS, Lechan RM. Leptin prevents fasting-induced suppression of prothyrotropin-releasing-hormone messenger ribonucleic acid in neurons of the hypothalamic paraventricular nucleus. Endocrinology 1997; 138:2569-2576.
Leibel R, Rosenbaum M. Changes in energy expenditure resulting from altered body weight. N Engl J Med 1995; 332:621-628.
Nillni EA, Vaslet C, Harris M, Hollenberg A, BjØrbaek, Flier J. Leptin regulates prothyrotropin-releasing hormone biosynthesis. Evidence for direct and indirect pathways. J Biol Chem 2000; 275(46):36124-36133.
Nillni E, Sevarino K. The biology of pro-thyrotropin-releasing hormone-derived peptides. Endocr Rev 1999; 20(5):599-648.
Palkovits M, Brownstein M. Maps and guide to microdissection of the rat brain. Elsevier, New York, 1988.
Van Haasteren GAC, Linkels E, Klootwijk W, van Toor H, Rondeel JMM, Temen APN, de Jong FH, Valentijn K, Vaudry H, Bauer K, Visser TJ, Greef WJ. Starvation-induced changes in the hypothalamic content of prothyrotrophin-releasing hormone (proTRH) mRNA and the hypothalamic release of proTRH-derived peptides:role of the adrenal gland. Journal of Endocrinology 1995; 145:143-153.
Watts AG, Sánchez-Watts G, Kelly AB. Distinct patterns of neuropeptide gene expression in the lateral hypothalamic area and arcuate nucleus are associated with dehydration-induced anorexia. J Neurosci 1999; 19(14):6111-6121.
Yen P. Physiological and molecular basis of thyroid hormone action. Physiol Rev 2001; 81(13):1097-1142.
Tags: anorexia y, la anorexia, anorexia, diferencial, hipotálamohipófisistiroideo, regulación
- GENERAL MESSAGE TO POSITION FROM POSITION SUBJECT DATE TIME
- VYŠŠÍ ODBORNÁ ŠKOLA EKONOMICKÁ ZLÍN BAKALÁŘSKÉ STUDIJNÍ PROGRAMY
- INSTRUCCIONES DE DILIGENCIAMIENTO PLANILLA DE RECIBO DE PAGARES Y
- ANNUAL DECLARATION FORM 20092010 CARRIAGE OF HEAVY FUEL OIL
- LEY PARA EXENCIÓN DE IMPUESTOS A ASOCIACIONES DEPORTIVAS SOCIALES
- NZQA REGISTERED UNIT STANDARD 29209 VERSION 1 PAGE 4
- SYLLABUS I NASTAVNI PLAN PREDMETA NA PREDDIPLOMSKOM I
- GETTING STARTED SELLING TO THE GOVERNMENT OF CANADA PREPARED
- 18 FIGUEROA ILLANES HUIDOBRO SALAMANCA ESTUDIO DE ABOGADOS COMMITTEE
- ASSEMBLY NO 2984 STATE OF NEW JERSEY 212TH LEGISLATURE
- MANCOMUNIDAD DE MUNICIPIOS DE LA COSTA DEL SOL OCCIDENTAL
- II AZ EMBERI ERŐFORRÁSOK MINISZTÉRIUMA ÁLTAL MEGHIRDETETT ÉS TÁMOGATOTT
- VILKET L NAMN KLASS ÖVNINGAR 1 VILKEN
- ¿QUÉ ES EL ENFOQUE CENTRADO EN LA PERSONA? (
- DECEMBER 21 2004 INTERNATIONAL FINANCIAL INSTITUTIONS INTERNATIONAL CONTRACTORS’
- F ACULTAD DE ARQUITECTURA Y URBANISMO SÍLABO INFORMACIÓN GENERAL
- 67 AÑOS A TU LADO ASOCIACIÓN ESPAÑOLA CONTRA EL
- WOGA274 PÁGINA 3 OMPI S WOGA274 ORIGINAL INGLÉS FECHA
- DAVID RIAZANOVS KARL MARX AND FREDERICK ENGELS AN
- ELEVEN PROVEN TECHNIQUES DESIGNED TO HELP YOU BECOME DEPRESSED
- CONVENIO DE SUBVENCIÓN ERASMUS+ EDUCACIÓN SUPERIOR PARA ESTUDIOS CONVENIO
- «УТВЕРЖДАЮ» ФИО МП «08» ДЕКАБРЯ 2020 ПРОТОКОЛ №232721 О
- FORENINGEN TIL STØTTE AF SYGDOMSRAMTE ERHVERVSAKTIVE BORGERE ERHVERVSAKTIVE MED
- FUNCTIONAL ANATOMY MUSCULOSKELETAL ANATOMY KINESIOLOGY AND PALPATION FOR MANUAL
- PEJ02 NORMAS PARA LA TRAMITACIÓN DE LOS MODELOS PEJ01
- ESTADÍSTICA DESCRIPTIVA PRÁCTICAS TEMA 1 CURSO 20052006 FACULTAD DE
- COMMUNICATING WITH APPLICANTS OBJECTIVES THIS GUIDE EXPLAINS THE
- МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ
- INFORME DE LA VISITA DE LAS EXPERTAS CUBANAS BERTA
- “RECONOCER INTERPRETAR ELEGIR” UN ESTILO DE PASTORAL EN ESTA
UZASADNIENIE DO PROJEKTU UCHWAŁY RADY GMINY ŁUBNIANY Z DNIA
LEKCJA 4 (29102020) TEMAT PORÓWNYWANIE LICZB NATURALNYCH – ĆWICZENIA
22 NÁRODNÍ CENTRUM PODPORY TRANSFORMACE SOCIÁLNÍCH SLUŽEB NÁRODNÍ CENTRUM
DIA DE BENIDORM 2017 COMPANYS I COMPANYES DE CORPORACIÓ
………………… DNIA WNIOSKODAWCA WNIOSKODAWCY IMIĘ I NAZWISKO…………………………………………………………
 Scenariusz Zajęć Rozwijających Kompetencje Emocjonalno Społeczne „z Uczuciami
Scenariusz Zajęć Rozwijających Kompetencje Emocjonalno Społeczne „z UczuciamiUMOWA NAJMU (ZWANA DALEJ „UMOWĄ”) ZAWARTA W DNIU
 APPEAL NUMBER OA165262010 UPPER TIER TRIBUNAL (IMMIGRATION AND ASYLUM
APPEAL NUMBER OA165262010 UPPER TIER TRIBUNAL (IMMIGRATION AND ASYLUMHUSITSTVÍ HUSITSTVÍ VYPLŇUJE V ČESKÝCH DĚJINÁCH VÍCE NEŽ JEDNO
1 PYLORUS STENOSIS KÉPALKOTÓ VIZSGÁLATA ULTRAHANG 2 AZ INVAGINATIO
 1272021 0 DRAFT 1 CBD CONVENTION ON BIOLOGICAL DIVERSITY
1272021 0 DRAFT 1 CBD CONVENTION ON BIOLOGICAL DIVERSITYPROPOSAL BUDGET CHECKLIST ALL PROPOSAL BUDGETS MUST BE REVIEWED
 MCPFE DATA COLLECTION FOR THE REPORT ON THE STATE
MCPFE DATA COLLECTION FOR THE REPORT ON THE STATE HEMOVET ® ÁLLATORVOSI KLINIKAI LABORATÓRIUM 1133 BUDAPEST THURZÓ
HEMOVET ® ÁLLATORVOSI KLINIKAI LABORATÓRIUM 1133 BUDAPEST THURZÓ UMOWA NR OG 35262018 ZAŁ NR 2 ZAWARTA W
UMOWA NR OG 35262018 ZAŁ NR 2 ZAWARTA WLA DOCUMENTAZIONE DEVE ESSERE PRESENTATA IN FORMATO CARTACEO ANCHE
 a Zala Megyei Közgyűlés Hatályos Rendeletei 2020 Június 2
a Zala Megyei Közgyűlés Hatályos Rendeletei 2020 Június 2VERTEGENWOORDIGING UITOEFENING VAN HET BEWIND OVER DE PERSOON PERIODIEK
 T C KONYA TEKNİK ÜNİVERSİTESİ LİSANSÜSTÜ EĞİTİM ENSTİTÜSÜ TEZ
T C KONYA TEKNİK ÜNİVERSİTESİ LİSANSÜSTÜ EĞİTİM ENSTİTÜSÜ TEZANEXA 3 LA NORMELE DE APLICARE A LEGII DREPTURILOR
